A-7 Ranch, Bellota – Redington Pass area
Arid Lands Resource Sciences 642
Spring 2000
USE AND MANAGEMENT OF ARID LANDS
Adapting land use to climate variability in arid lands:
Ranching and the concept of grass banks in southern Arizona
IV. Vegetation, Fire, and Climate History
Kathryn Mauz, Rick Krenzer, and Luis Cervera
4.1. INTRODUCTION
Elements of the physical environment vary with time and across space, structuring, to some extent in the immediate term and to a large extent ultimately, human land use practices. Annual distribution of rainfall, periodic and enduring drought, landscape disturbance, and vegetation change underlie, and in some cases are outcomes of, range management decision making. The present review introduces aspects of the physical environment - climatic variability and drought, vegetation ecology and vegetation change, and fire ecology and history of fire - that may be important to implementing a "grass bank" management system on a semidesert rangeland. Discussion draws on range management literature for southeast Arizona, and attempts to apply these findings to the A-7 Ranch on the west side of the San Pedro River Valley, east of Tucson.
4.2. THE PHYSICAL GEOGRAPHIC SETTING OF THE A-7 RANCH
The study area that includes the A-7 Ranch occupies T.12-13 S and R.17-19 E (Gila Principal Meridian and Baseline), straddling the boundary between Pima County and Cochise County in southeastern Arizona. The area is situated between Redington Pass on the west at about 4000 feet (1220 m) elevation, and the San Pedro River on the east at about 3000 feet (920 m) elevation; Buehman Canyon runs along the north end and Roble Canyon cuts across on the southeast. The southwestern boundary of the A-7 property is shared with the Coronado National Forest.
Redington Pass occupies a middle Tertiary-age mylonite zone associated with the Santa Catalina-Rincon metamorphic core complex. Four granitic intrusives are associated with the Catalina (north) half of the complex: the Precambrian Oracle Granite and the Tertiary plutons Leatherwood Quartz Diorite, Wilderness Granite, and Catalina Granite. Within the study area, the Oracle Granite is exposed south and east of Piety Hill, while the Wilderness Granite is exposed at and north of Piety Hill; small exposures of the Precambrian Pinal Schist (deformed basement rocks) may also occur at the eastern edge of the Wilderness Granite (Naruk & Bykerk-Kauffman, 1990). Most of the Bellota grazing allotment on the Coronado National Forest is underlain by the Wilderness Granite, the erosion of which contributes to the uneven, knob-and-swale topography supporting the oak-savanna (encinal) vegetation community.
The intruded countryrock includes Paleozoic through Cretaceous sedimentary rocks that exhibit varying degrees of metamorphism and deformation (Schloderer, 1974). These rocks may include low-grade metamorphic rocks (sandstones, limestone- and chert-cobble conglomerates) of the Lower Cretaceous Bisbee Group (Naruk & Bykerk-Kauffman, 1990) and Mississippian-Permian strata that include the medium- to coarse-grained, gray, Mississippian Escabrosa Limestone and a thin package of metamorphosed sedimentary rocks collectively known as the Permian Horquilla Limestone (Schloderer, 1974). These rocks are exposed northeast of Piety Hill at the northeastern boundary of the Wilderness Granite, along the Redington Pass Road and in a canyon immediately to the north of and below the road on the east side of Redington Pass (Naruk & Bykerk-Kauffman, 1990). Vegetation may be locally unique on these substrates, particularly where surface water is available in canyon bottoms (see Malusa & Porter, 1990, for example).
The majority of the area occupied by the A-7 Ranch lies east of Redington Pass on tilted surfaces of deformed, variably cemented, Pliocene-age gravels descending from the Catalina-Rincon mountain front, and on younger (late Pliocene-Quaternary) valley fill and terraces of the San Pedro River (Zimmermann, 1969). It is on these surfaces that the semidesert grassland and Sonoran upland vegetation communities occur in the study area.
4.3. HISTORY OF CLIMATIC VARIABILITY IN SOUTHEASTERN ARIZONA
4.3.1. Precipitation
Seasonal and Annual Patterns
Southeastern Arizona's climate is semiarid. Average annual rainfall ranges from 9 to 25 inches (229 to 635 mm). The rainfall pattern in southeastern Arizona is bimodal. Summer precipitation during July, August, and September ranges form less than 50% of the annual total in the northwestern part of the area to more than 70% in the southeastern part. The period of summer precipitation (known as the summer "monsoon") is quite sharply defined, beginning around the last week of June and lasting until the middle of September. Precipitation mainly occurs from convective cells initiated by surface heating, convergence or, less commonly, orographic lifting. These summer convective storms form in mesoscale clusters (Barry and Chorley, 1998).
Total annual precipitation in southeast Arizona has ranged from 193 to 636 mm, and averages 363 mm for the period 1895-1999. This total annual precipitation is shown in Figure 4-1. During the same period, winter (D-J-F-M) precipitation has ranged from 14 to 283 mm, and averages 102 mm. Summer (J-A-S) precipitation has ranged from 108 to 346 mm, and averages 194 mm (CLIMVISb). The average monthly precipitation and average monthly temperature for the Southeast Arizona climate division are shown in Figure 4-2.
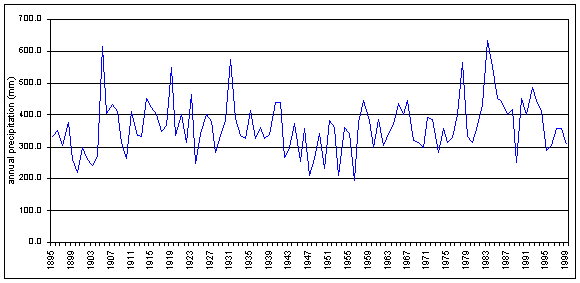
Figure 4-1. Total annual precipitation, Southeast Arizona, 1895-1999 (Data:CLIMVISb).

Figure 4-2. Climograph for Southeast Arizona (climate division 7), 1895-1999 (Data:CLIMVISa,b).
Within Arizona state climate division 7, seven stations within 100 km of and sharing features of their geographic settings with Bellota Ranch reveal the spatial, as well as interannual, variability of rainfall inherent to this area. Data sets for these stations - Tucson, Oracle, Fort Grant, Willcox, Tombstone, Bowie, and Safford - span 98 to 134 years up to 1989 (CLIMVISb). Average annual precipitation for these seven stations ranges from 230 to 515 mm, and the average for all stations over all years is 334 mm. Average winter (D-J-F-M) precipitation for these stations ranges from 70 to 201 mm. The average winter precipitation for seven stations over all years is 104 mm, an average of 30% of the annual total. Average summer (J-A-S) precipitation ranges from 116 to 214 mm. The average summer precipitation for seven stations over all years is 164 mm, an average of 50% of the annual total. Seasonal and annual precipitation for the seven weather stations around the A-7 Ranch are graphed in Figure 4-3.
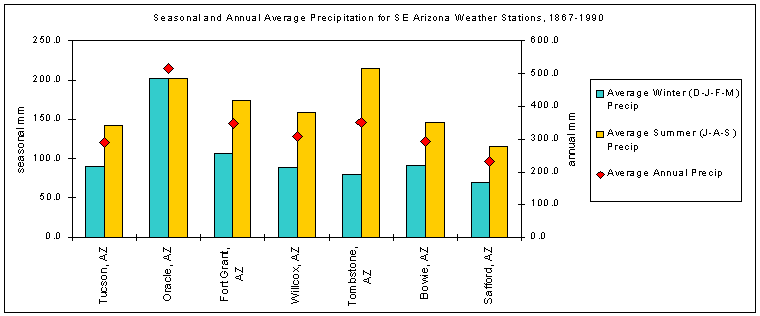
Figure 4-3. Seasonal and annual precipitation for seven weather stations in Arizona climate division 7
nearest to and surrounding the A-7 Ranch (Data:CLIMVISb).
Annual precipitation for an independently maintained station on the Bellota Ranch for the period 1981 to 1997, ranges from 316 to 784 mm and averages 478 mm. Winter precipitation (D-J-F-M) ranges from 56 to 348 mm, and averages 181 mm. Winter precipitation averages 38% of annual rainfall at this location. Summer precipitation (J-A-S) on the Bellota Ranch ranges from 100 to 273 mm and averages 194 mm. On average, summer precipitation represents the 41% of the annual total. The average monthly precipitation for the Bellota Ranch is shown in Figure 4-4.
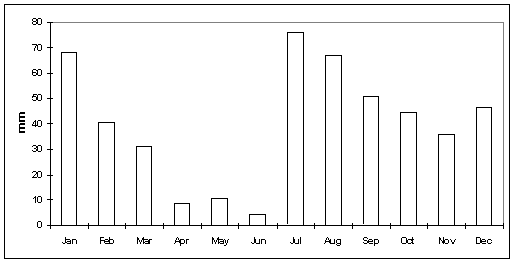
Figure 4-4. Average monthly precipitation, Bellota Ranch, Arizona, 1981-1997.
The El Niño-Southern Oscillation
The El Niño phase of ENSO tends to displace the winter storm track farther south, placing Arizona and the Southwest in the path of a greater number of more intense storms. Winter (D-J-F-M) precipitation in southeast Arizona during an El Niño winter averages 7.0 inches (177.8 mm), nearly twice the 102-year historical average for the same months (CPC, 2000b). Historically, strong El Niño episodes have been characterized by above normal winter precipitation statewide, with totals in northern Arizona averaging 140% of normal and totals in the southern part of the state averaging up to 180% of normal (CPC, 1997). Winter precipitation during El Niño may be 50-75% higher than when La Niña conditions prevail (Brown, 1999). Likewise, temperature during an El Niño winter is colder by an average of 1.2 degrees Fahrenheit than the long-term average winter temperature of 54.3 oF (12.4 °C) (CPC, 2000c).
Historic Trends
Between 1898 and 1960, southern Arizona experienced a slight decrease in rainfall together with a slight increase in temperature (Sellers, 1960). The pattern of change includes both an increase in light rainfall and a decrease in intense rainfall events, however there have not been statistically significant changes in the temporal distribution of rainfall during this period (Bahre, 1996). Over the next 100 years, precipitation in Arizona is estimated to decrease slightly in summer (with a range of 0 to -15%), to increase by 20% in spring (with a range of 10-40%), to increase by 30% in fall (with a range of 10-50%) and to increase by 60% (with a range of 30-100%) in winter (EPA, 1998).
4.3.2. Temperature
Seasonal and Annual Patterns
Average annual temperatures in southeast Arizona for the period 1895-1999 have ranged from 15.2°C to 19.1°C. Winter (D-J-F-M) temperatures for that same period averaged 9.3±1.0°C, while summertime (J-A-S) temperatures averaged 25.3±0.8°C. The average annual temperature in Southeast Arizona, over this time period is shown in Figure 4-5.
The average annual temperature at the Bellota farm was 19.3 °C for the period 1993-1997. Winter (D-J-F-M) temperatures for that same period ranged from 9.4°C to 14.4°C and averaged 11.4°C. Summer (J-A-S) temperatures ranged from 22.8°C to 29.7°C, averaging 27.1°C.
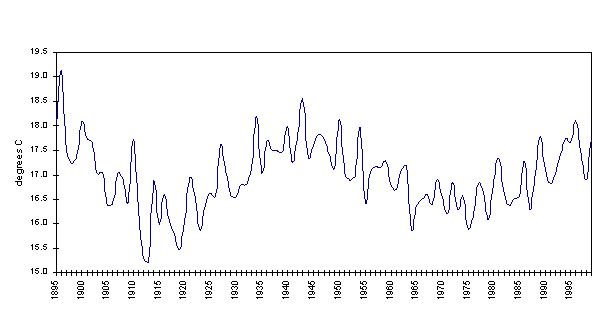
Figure 4-5. Average annual temperature, Southeast Arizona, 1895-1999 (Data:CLIMVISa).
The average temperature in Tucson, Arizona, has increased by 3.6 °F in the last century, and precipitation has increased by up to 20% in many areas of the state. Recent modelling efforts suggest that both the intensity of storm events and the frequency of extreme temperature events in Arizona could increase in the next century (EPA, 1998).
4.3.3. Drought
Because normal precipitation over most of the state is low in comparison with the very high potential evapotranspiration, drought is recognized as a frequently occurring phenomenon in Arizona (Steila, 1972). The pattern of drought is complex and can vary in intensity, frequency, and duration from year to year. The Palmer Drought Severity Index (PDSI) uses temperature and rainfall data in a formula to determine dryness, providing a measure of drought relative to normal local or regional conditions (Drought Information Center, 2000). PDSI is often used to monitor or predict irrigation water supplies, reservoir levels, range conditions, amount of stock water, and potential intensity of forest fires (CPC, 2000a).
Monthly PDSI value for Southeast Arizona vary on an interannual basis, from lowest in the winter months to highest in the summer months. Drought conditions are most common in Arizona in May and June (Sellers and Hill, 1974). Figure 4-6 illustrates the average monthly PDSI for southeast Arizona from 1895 to 1999. The average annual PDSI for this area and time period is plotted in Figure 4-7.
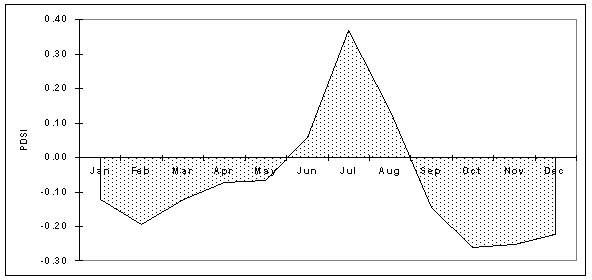
Figure 4-6. Average monthly PDSI, Southeast Arizona, 1895-1999 (Data:CLIMVISc).
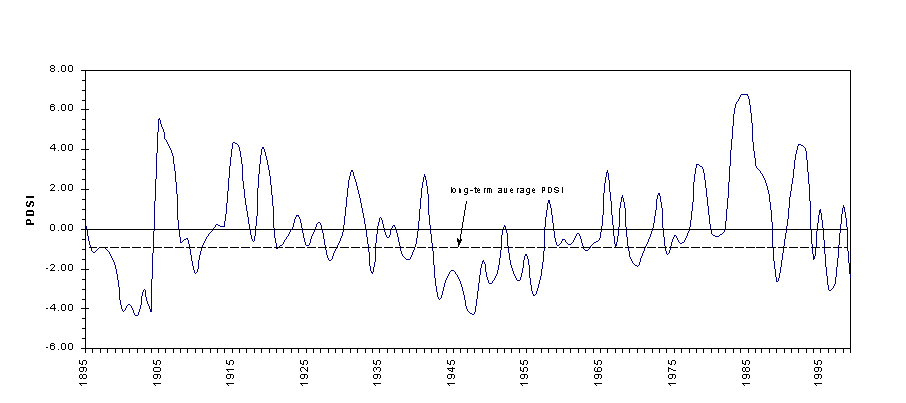
Figure 4-7. Average annual PDSI, Southeast Arizona 1895-1999 (Data:CLIMVISc).
Steila (1972) devised an 11-class division of wet and dry conditions from which six classes of relative drought have been created for this study (Table 4-1). Within this classification, climate in southeast Arizona has been characterized by mild drought or incipient dry conditions in more than half of the years of the past century (1895-1999) (Figure 4-8). Years in which El Niño conditions prevailed in winter all fall within the two wettest PDSI classes, while all of the years in which La Niña conditions predominated fall in PDSI class 5. Despite this qualitative relationship between ENSO years and PDSI class, the Southern Oscillation Index (SOI) and PDSI data exhibit only a weak negative correlation with one another (r=-.14).
Table 4-1. Moisture anomaly index used to classify PDSI values.
|
Moisture anomaly index* |
Class |
Classification used in this work. |
|
> 4.0 |
Extremely wet |
(1) Very wet to extremely wet |
|
3.00 to 3.99 |
Very wet |
|
|
2.00 to 2.99 |
Moderately wet |
(2) Slightly wet to moderately wet |
|
1.00 to 1.99 |
Slightly wet |
|
|
.50 to .99 |
Incipient wet spell |
(3) Near normal to incipient wet spell |
|
-0.49 to 0.49 |
Near normal |
|
|
-0.50 to - 0.99 |
Incipient dry spell |
(4) Mild drought to incipient dry spell |
|
-1.00 to -0.99 |
Mild drought |
|
|
-2.00 to -2.99 |
Moderate drought |
(5) Moderate drought to severe drought |
|
-3.00 to -3.99 |
Severe drought |
|
|
< -4.0 |
Extreme drought |
(6) Extreme drought |
|
* Source: Steila, 1972 |
||
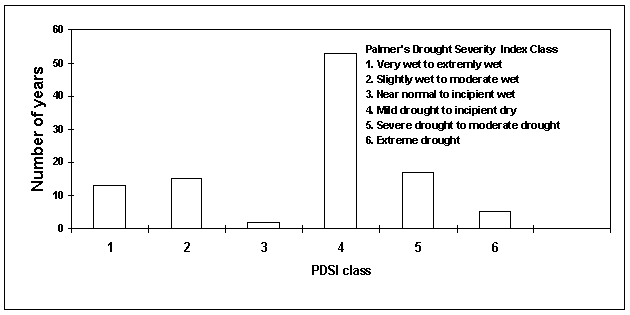
Figure 4-8. Frequency of years within PDSI classes, southeast Arizona, 1895-1999.
Streamflow data for the San Pedro River at Redington, Arizona, were obtained from the U.S. Geological Survey data sets (USGS, 2000). The average annual flow between 1950-1997 is 43.3 cubic feet per second (cfs), ranging from 0.6 cfs in 1997 to 177.4 cfs in 1955. A decrease in the average annual discharge is well defined in the 20-year period from 1955 to 1975 (Figure 4-9). The annual average streamflow exhibits a weak positive correlation with the Palmer Drought Severity Index (r=0.30 at P=.05). In general, increased average streamflow corresponds to positive PDSI values (wet periods), while decreased average streamflow corresponds to negative PDSI values (dry periods).
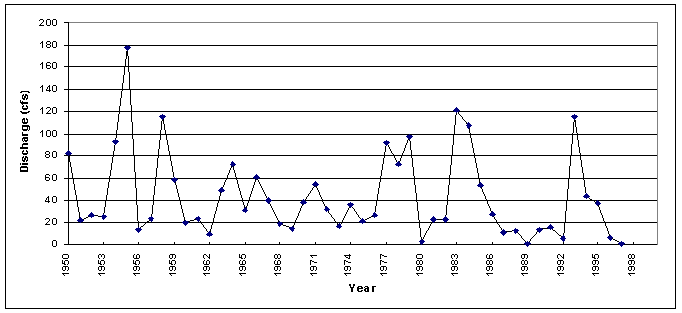
Figure 4-9. Annual average stream flow at Redington, Arizona, 1950-1997 (Data:USGS, 2000).
4.3.4. Implications of Climate Change for Range Management
Climate change can have implications for both natural and managed grassland ecosystems. Livestock production may remain unaffected unless summer temperatures rise significantly and conditions become significantly drier. Under these conditions, livestock tend to gain less weight and pasture yields decline (EPA, 1998).
Runoff in Arizona primarily results from summer thunderstorms and winter precipitation. In the mountains, most of the winter precipitation falls as snow. A warmer climate could mean less winter snowfall, more winter rain, and a faster, earlier snowmelt. Higher temperatures and increased evaporation could lower lake levels and streamflows in the summer (EPA, 1998).
Hotter, drier weather could increase the frequency and intensity of wildfires. Expansion of desert into former rangeland or of woodland into grassland could also be consequences of climate change with implications for the livestock industry (EPA, 1998). The effects of precipitation and temperature changes on range resources will vary depending on the relative change in timing, intensity, and duration of extreme events within and between years.
4.4. SEMIARID GRASSLAND AND OAK-SAVANNA VEGETATION IN SOUTHEASTERN ARIZONA
4.4.1. Regional Biogeographic Setting
The semidesert grassland in southeast Arizona is contiguous with the Chihuahuan desert and is probably allied more with the Chihuahuan than the Sonoran biome, although this community may be found with Sonoran Desert taxa at its western limits (Brown, 1994). At a regional scale, the Redington Pass area and A-7 Ranch occur near the northwestern limit of this vegetation community. Here, the semidesert grassland occurs between about 1100 m and 1400 m (3600-4600 feet) elevation (Brown, 1994), with transitions to Madrean Woodland above in the foothills of the Santa Catalina and Rincon Mountains and to Arizona Upland and Chihuahuan Desertscrub vegetation communities below in the lower San Pedro River valley. These transitions may be complex, reflecting elevational, spatial, topographic, and edaphic gradients underlain and structured by Basin-and-Range bedrock and surficial geology, and local and regional climatic patterns.
4.4.2. Vegetation Communities in the Area of the A-7 Ranch
Regional-scale vegetation maps that include southeastern Arizona have been developed by Brown and Lowe (1994), the USDA Natural Resource Conservation Service (NRCS), and as part of the Arizona Gap Analysis Project (GAP) (see Hobbs, 1999). The vegetation types identified for the area including the A-7 Ranch are summarized in Table 4-2. A portion of the NRCS Major Land Resource Areas map is included as Figure 4-10.
Table 4-2. Correlation of vegetation communities for the area of the A-7 Ranch as defined in three regional vegetation maps.
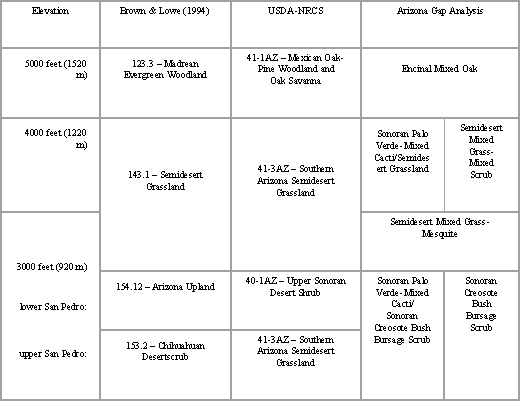
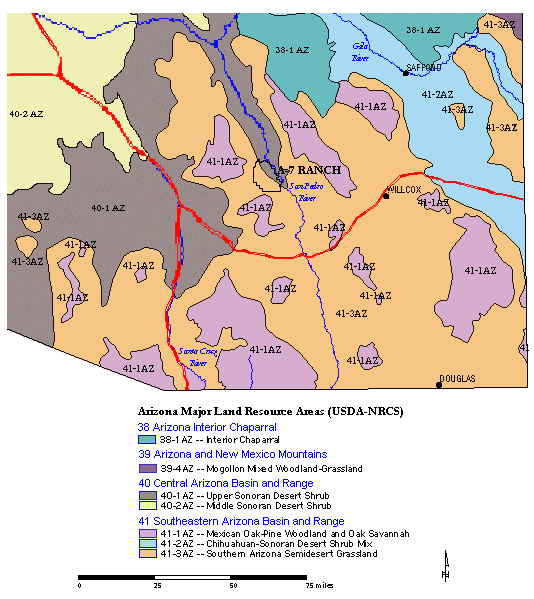
Figure 4-10. USDA-NRCS Major Land Resource Areas in southeast Arizona,with the location of the A-7 Ranch indicated (NRCS,1999).
Quimby (1976) reported four species of grama grasses (Bouteloua chondrosioides, B. hirsuta, B. curtipendula, B. filiformis) as the primary forage grasses on the Bellota grazing allotment of the Coronado National Forest, an area of about 36,000 acres that straddles Redington Pass and borders the A-7 ranch on the west. Other important forage plants on the allotment at that time included fairy duster (Calliandra eriophylla), ratany (Krameria spp.), and shrubby buckwheat (Eriogonum wrightii), along with minor amounts of several other grasses (Quimby, 1976). Scrub-shrub taxa are diverse and, along with both stem and leaf succulents, are present in all areas of the A-7 Ranch. Arboreal taxa, notably velvet mesquite (Prosopis velutina) may be common, as well, and oak-savanna vegetation (NRCS Oak-Savanna and GAP Encinal Mixed Oak) may occur as the upper elevation transition to Brown's (1994) Madrean Woodland.
In the Redington Pass area, oak-savanna species include oaks (Quercus oblongifolia, Quercus emoryi) and juniper (Juniperus sp.), along with several chaparral-shrub taxa (e.g., Arctostaphylos pungens, Garrya wrightii) that become more common at higher elevations (see Figure 4-11b). Leaf succulents at upper elevations on the Bellota Ranch include Agave schottii, Agave palmeri, Nolina microcarpa, Dasylirion
wheeleri, and Yucca sp. Descending into semidesert grassland and Sonoran-Chihuahuan desertscrub communities of the San Pedro River valley (see Figure 4-11a), creosote bush (Larrea divaricata subsp. tridentata)
becomes ubiquitous, ocotillo (Fouquieria splendens) and soaptree yucca (Yucca elata) appear, and saguaro (Carnegiea gigantea) is increasingly common, particularly on alluvial surfaces, in the grassland. Woody vegetation
of wash margins, seeps, and occasionally swales may include the shrubs Acacia constricta, Acacia greggii, Zizyphus obtusifolia, and Celtis pallida, the trees Cercidium floridum and Cercidium microphyllum (at mid- to lower elevations), Celtis reticulata (all areas), and, uncommonly, broad-leaf deciduous trees (e.g., Populus sp.) in well-watered drainages and at lower elevations.


(a) Sonoran and Chihuahuan Desert scrub.

(b) Oak-juniper savanna.

(c) Canyon floor riparian areas.
Figure 4-11 (a-c). Vegetation communities on the A-7 Ranch.
Malusa and Porter (1990) surveyed canyon vegetation on and peripheral to the A-7 ranch (e.g., Figure 4-11c). The authors identified Arizona sycamore (Platanus wrightii), velvet ash (Fraxinus velutina), Goodding's willow (Salix gooddingii), Arizona walnut (Juglans major), and Fremont's cottonwood (Populus fremontii) along Buehman Canyon. Cottonwood, ash, and walnut were observed in Youtcy and Espiritu Canyons, and willow, along with cattail (Typha domingensis), were noted in Youtcy Canyon. The canyon bottom in Espiritu Canyon is described as supporting more xeroriparian species, including burrobush (Hymenoclea salsola), desert broom (Baccharis sarothroides), and, on hillsides, jojoba (Simmondsia chinensis). Roble Canyon has relatively fewer cottonwood and ash, but is notable for other riparian vegetation including maiden's hair fern (Adiantum cappilus-veneris). The authors emphasize that these riparian areas provide important habitat for several rare and endangered plants and animals, and serve as nurseries for young trees and sources of seed to support future recruitment. As such, potentially damaging land uses (e.g., mining, cattle grazing) should be minimized or eliminated in these canyons (Malusa and Porter, 1990).
4.4.3. Semidesert Grassland-Oak Savanna Range Plant Phenology and Response to Drought
Oak-savanna vegetation tends to occupy areas of moderate elevation that are more mesic than valley floors yet more temperate than higher elevations. In a study of oak-savanna vegetation ecology in southeastern Arizona, Kincaid (1959) found that this community is most well-defined between 1370 m and 1980 m (4000-6500 feet) elevation, and transitions to other vegetation types at the upper and lower limits reflect a general elevational gradient in precipitation and temperature. Shortgrasses (e.g., Erioneuron pulchellum) found in these areas tend to occur near the lower boundary of the oak-savanna and be more tolerant of lower-moisture conditions. Midgrasses (e.g., Andropogon sp., Muhlenbergia spp., Bouteloua spp., Aristida spp.) tend to occur at higher elevations in the oak-savanna where moisture levels tend to be higher, and different species of midgrasses tend to be distributed preferentially with elevation within this category (Kincaid, 1959).
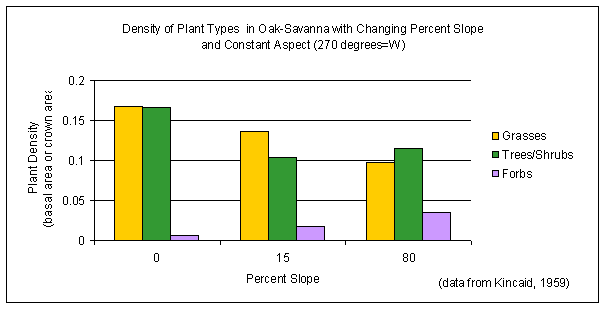
Both the spatial distributions and densities of characteristic plant types in the oak-savanna have been found to vary with
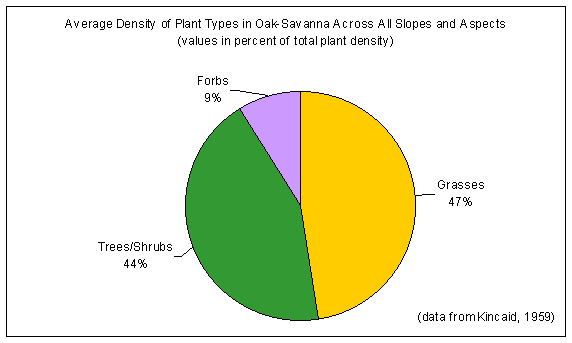
slope and aspect, as well as with elevation. In study plots having the same exposure (west-facing slopes), grass density was found to decline with increasing percent slope (Figure 4-13a). Across all slopes and aspects (Figure 4-13b), grasses were the dominant constituents of the oak-savanna (47% of total plant density by basal area), with trees and shrubs about the same (44% of total plant density by crown area) and forbs occurring as minor constituents (9% of total plant density by basal area) of the community (Kincaid, 1959).
|
|
|
|
|
Figure 4-13. Composition of southeast Arizona oak-savanna vegetation. |
The timing of vegetation growth and amount of production, and the variability in productivity across space, in the semidesert grassland is influenced by annually low and seasonally variable (but always bimodal) rainfall, high evapotranspiration, plant physiology (particularly water uptake and/or storage strategies), and soil characteristics (Haile, 1981; McClaran, 1995; Burgess, 1995). In a study of the energy flow dynamics of a perennial-grass ecosystem in northern Arizona, Davey (1980) concluded that moisture is the primary limiting factor of grass productivity. He found that, although C4 (warm-season) grasses may produce vegetative growth in the spring, they produce significantly more in the summer (Davey, 1980). Other research in the grasslands of southeast Arizona suggests the same, and emphasizes the importance of summer growth (about 90% of annual grass productivity) over relatively insignificant winter growth (10% of annual grass productivity) for species in this area (McClaran, 1995).
Particularly in the warm Southwestern deserts, despite increased precipitation during the summer months, unusually high summer temperatures may be limiting due to relatively high rates of evapotranspiration. Prolonged drought in grassland vegetation can have negative consequences both for productivity of existing plants and for propagation and stand replacement. Davey (1980) found that reproduction of perennial grasses by seed is insignificant during low-rainfall years. Based on an additional finding that summer growth relies upon exceeding "a specific soil moisture threshold in the top 10 cm ... for approximately three days" and a simulation analysis of grass dynamics, he concluded that, "at annual precipitation levels below 10-12 cm, grass production is substantially reduced with relatively small decreases in annual precipitation" (Davey, 1980:135-136). Records for permanent transects monitored on the Bellota grazing allotment on the Coronado National Forest indicate that the 2-½ year drought in 1954-1956 "devastated" the range there (Quimby, 1976:12).
4.4.4. Grazing Impacts to Grass Plants
Brady (1995) identifies grazing and fire suppression as two physical environmental stressors having potentially broad and severe ecological impacts in Arizona's grasslands, including the semidesert grasslands in the southeastern part of the state. Grass plants are impacted by herbivory through removal of new leaves and canopy, exposure of older/basal leaves, modification of soil chemistry, texture, and hydrodynamics, and grazing selectivity. These processes can result in diminished productivity and changes to the composition of range vegetation. There may be both short-term and long-term consequences for forage resistance to stress, resilience following the removal of stress, and restorability of a range after it has been physically changed due to stresses associated with overgrazing (Brady, 1995).
Several factors have been cited by other authors that could affect the relationship between forage value and grazing impacts to forage plants. These may include 1) spatial heterogeneity of pasture resources and physical attributes such as soils, 2) the duration of grazing, 3) the timing of grazing relative to the grass life cycle and annual variation in temperature and precipitation, 4) the individual grass species involved, 5) feedback dynamics between grazing and grazed plants, and between species of grazed plants in the same pasture (see Agnew, 1997), and 6) the degree to which supplementary feeding and watering of livestock is undertaken, including the number and location or spacing of water supplies and supplemental resources such as salt. Hodgkinson and Mott (1987) suggest that the responses of a pasture to grazing may differ depending on the phenological stage and prevailing environmental conditions at the time that grazing impacts occur: "...for much of the time the physiological state of grasses is such that they cope with the level of grazing intensity imposed by the conservative grazing systems. Only when the grazing intensity exceeds a threshold level during a period of prolonged low or high soil moisture or a stage of phenological development, are grasses and pastures at risk of impaired growth and survival" (Hodgkinson & Mott, 1987:186). Alterations to soils caused by grazing may include compaction, depletion of soil nutrients, and removal of
topsoil. These, in turn, influence water infiltration as well as limit the kinds of plants that can become established on a site.
4.4.5. Consequences for Vegetation Communities
"Perennial grasses...are the dominant element in most sown and natural pastures, they usually constitute the major component of the diet selected by grazing animals, and their retention is considered essential for the stability of managed grazing systems" (Hodgkinson & Mott, 1987:172). Studies on grazed and conserved areas in desert grassland vegetation on the Page Ranch north of Oracle Junction reveal diminished species richness and soil fertility on grazed portions. Further, vegetation on grazed portions was predominantly shrubs whereas on conserved areas perennial grasses were most abundant (Haskell, 1945). Kincaid hypothesized that the sensitivity of midgrasses to slight changes in soil moisture, combined with the impacts to soil microclimate by cattle grazing noted by other authors, could lead to declines in midgrasses and increases in shortgrasses independent of the impacts to the plants themselves by grazing animals. In undisturbed oak-savanna, midgrasses were found to be the dominant grasses in the community, whereas on grazed areas the proportion of shortgrasses was higher (Kincaid, 1959).
Vegetation changes on the Bellota allotment on the Coronado National Forest have been attributed to overstocking and overgrazing, as well as to improper placement of salt and lack of supervision of cows on the range through the middle decades of the 20th century. "A review of the last four Production and Utilization Studies (1968, 1971, 1973 and 1974) conducted on the Bellota Allotment as well as a comparison of the current [1976] range environmental reanalysis with the previous analysis indicate a worsening trend in Production and Utilization of Forage Plants and a decrease in range condition. ... The average pounds production per acre has been quite low and reflects the past overgrazing practices" (Quimby, 1976:4). Quimby (1976) noted of riparian vegetation in canyons on the Bellota grazing allotment that "excessive livestock grazing has eliminated almost all reproduction of these trees and the stands are now either mature or approaching a decadent state. If overuse continues, it is only a matter of time before these trees will be gone along with their dependent wildlife" (Quimby, 1976:5). Of browse species, Quimby describes a pattern of excessive "hedging," indicating repeated pruning by cattle during the plants' growing seasons and failure for the new growth to persist. This pattern is suggested to have
consequences for browse-dependent wildlife as well as for persistence of browse plant populations (Quimby, 1976). Additionally, remediation of erosion due to vegetation losses in Redington Pass has been necessary in the past, and overgrazing-related vegetation changes (notably incursions of woody plants) have led to implementation of eradication experiments and ongoing programs by the Forest Service on the Bellota allotment.
4.4.6. Patterns of Vegetation Change in the Semidesert Grassland-Oak Savanna, Southeastern Arizona, in the 20th Century: A Brief Overview
Grasses
The "original" (i.e. pre-European), predominant vegetation of the semidesert grassland comprised perennial, summer-active bunch grasses, reproducing primarily from seed and forming a discontinuous cover on the landscape (Brown, 1994; Griffiths, 1900, in Haskell, 1945). Although widespread presence of grasses gives these ecosystems their name, other growth forms - subshrubs, shrubs, succulents, and some trees - are commonly found within grasslands and in varying abundances relative to the grasses. These occurrences reflect and respond to differences across space and changes over time in aspects of the physical environment (e.g., rainfall, soils, winter and/or summer temperatures) and in disturbance regimes (e.g., herbivory, fire) (Burgess, 1995). Differences in grassland composition and structure may occur patchily at varying scales, and changes may be manifest on the order of a few years to decades. Grassland species structure and composition in southeastern Arizona have been altered in many places by human land use practices - including cattle grazing, range seeding, fire suppression, and resulting changes in hydrology and soils - since the late nineteenth century. Typically, several grass species may be dominant, however native perennial bunch grasses may have been replaced in places by low, sod-forming grasses or by annual species or non-native bunch grasses (Brown, 1994; Haskell, 1945). In a former grassland area overgrazed by sheep and cattle in the early part of the 20th century, Haskell (1945) observed a higher frequency of all types of annual plants and lower frequencies of all types of perennials than on ungrazed plots. Haskell also documented shrub incursions, notably of burroweed and cacti, into heavily grazed plots in former grassland near Oracle, concomitant with the disappearance of the native perennial grasses. These observations suggest that the pattern of demographic change in grasses may extend to the entire vegetation community.
Woody Plants
In an early and often-cited manuscript, "Vegetational Changes on a Semidesert Grassland Range from 1858 to 1963," Buffington and Herbel (1965) report that large areas of the desert southwest have been "invaded" by indigenous woody plants since the middle 1800s. In the semidesert grasslands in southeastern Arizona, most of this change has taken place since the 1890s (Bahre and Shelton, 1993). Several factors are claimed to influence this pattern - climate change, overgrazing, fire suppression, changes in hydrology, changes to soils, changes in seed dispersal patterns. The influence of particular mechanisms is debated, and insofar as these are not independent, there are likely multiple, interacting processes responsible.
Repeat photography of permanent transects on the Bellota grazing allotment indicates mesquite establishment in the 1950s and
1960s, with plants growing to 1-3 feet tall in 6 years or less (Quimby, 1976). In the 1976 Bellota Range Environmental Analysis, 15% of the suitable range on the allotment was considered to have been invaded by velvet mesquite (Quimby, 1976). Other "invasive" woody plants on the Bellota grazing allotment include burroweed, turpentine bush, and wait-a-minute bush. The invasions of these shrubs, as well as of mesquite and amole (Figure 4-14, below), into former grassland are considered to have been facilitated by overgrazing and the consequent removal of fire-carrying fine fuel from the landscape (Quimby, 1976).
|
|
|
|
Figure 4-14. Some problematic woody plant species in Redington Pass. |
|
 (a) velvet mesquite
(a) velvet mesquite (b) turpentine bush
(b) turpentine bushOther Species
The leaf succulent, Agave schottii Engelm., commonly called 'amole' and 'shin dagger', grows at upper elevations in
semidesert grassland and oak savanna, often on coarse soils and rocky slopes. It can form dense stands that, particularly due to its rosette of stiff, incurved, upright leaves, can both be impenetrable to livestock and preclude the growth of forage species. Amole has been recognized as problematic by range managers with the Coronado National Forest since the 1950s. Repeat photography of permanent transects on the Bellota grazing allotment reveals that, while some populations of amole suffered severe damage in the 1954-1956 drought, the plants recovered and were again invasive by the 1970s (Quimby, 1976). The
Forest has combated amole with prescribed fire since at least as early as the 1960s. This policy has not been without contention, and its benefits are debated (Quimby, 1976). Amole that was burned is pictured regrowing in Figure 4-15, despite that the same blaze was sufficient to kill manzanita growing nearby.
|
|
|
Figure 4-15. Amole regrowing after a fire that killed nearby manzanita. |

4.5. FIRE'S EFFECTS ON SEMIARID DESERT ECOSYSTEMS
Wildfire was a significant force in maintaining desert grassland ecosystems of the southwestern United States prior to Anglo-American settlement. Wildfires ignited primarily by lightning and also set by humans were common in southeastern Arizona before the 1890s (Bahre, 1995; Bahre, 1996; Leopold, 1924). The exclusion of fire in conjunction with heavy grazing has encouraged the invasion of woody species into grasslands. Overgrazing of rangelands, drought conditions, and an active policy of fire suppression has resulted in the transition of former desert grasslands into desert shrublands throughout much of southeastern Arizona (Bahre, 1995; Bahre, 1996; Leopold, 1924; Wright, 1990). These processes may, in part, be responsible for the current vegetation patterns present on the A-7 Ranch.
4.5.1. Impacts to the Physical Environment
Fire, either wild or prescribed, in semidesert grasslands and shrublands has a wide range of effects on the vegetation, soils, water, and watershed resources. Fire characteristics, season of burning, and preburn and postburn environmental conditions create highly variable responses of vegetation to fire in desert ecosystems (Wright, 1990).
Hydrology
Fire can significantly alter watershed conditions through the removal of vegetation and litter. Burning on semidesert rangeland may increase water yield and storm water runoff. The amount of increase depends on the intensity and severity of burning, and the size and slope of the area burned. Hydrologic processes that are affected by fire include the interception of precipitation, infiltration, overland flow, and soil water storage. Removal of vegetation by fire results in increases in direct evaporation and decreases in both water retention by litter and debris, and transpiration through plants. Fire indirectly affects microclimate by causing greater air and soil temperature extremes. Increased surface runoff due to fire may diminish water quality through increased sedimentation and turbidity, increased stream temperatures, and increased concentrations of nutrients (Baker, 1990).
The A-7 Ranch lies entirely within the San Pedro River watershed. Several canyons transverse the property and drainage generally flows in a northeastern direction. The northeastern boundary of the ranch borders approximately 9 miles of the San Pedro River. Plant communities on the A-7 change as a function of elevation. The composition of plant communities shifts from semidesert mixed grass-mixed shrub at high elevations; to semidesert mixed grass-mesquite at moderate elevations; to Sonoran creosotebush-bursage scrub, palo verde mixed cacti, and Chihuahuan creosotebush-tarbush scrub at low elevations; to Sonoran riparian vegetation along the San Pedro River (Hobbs, 1999). A reduction in burning potential might be expected to correspond with a decline in elevation and changes in vegetation type. Lower elevation communities composed of desertscrub species and cacti with intermittent patches of bare soil are less susceptible to fire than mixed-grass shrub communities. These areas could act as a buffer to increased postburn runoff from higher elevations and could impede the spread of wildfire to areas in which burning would not be desirable (e.g., riparian communities).
Soils
Leopold (1924) observed widespread soil erosion in southern Arizona, particularly along watercourses. He asserts that in spite of periodic fires, native grasslands prevented erosion and that overgrazing in the late 1800s and early 1900s caused heavy losses of soil from Arizona rangeland.
Erosion by wind, water, or gravity usually increases following fire. Biggs and others (n.d.) reported increased rates of surface runoff and erosion on burned sites compared with those of an unburned site. Large areas cleared by fire are vulnerable to erosion and can yield substantial amounts of eroded material if subjected to high-intensity summer storms immediately following burning. Lower-intensity winter rains often result in less erosion (Simanton et al., 1990). With the reestablishment of vegetative cover, and depending upon grazing intensity, erosion losses revert to preburn levels within a few years after burning (Baker, 1990).
Desert rangeland supports relatively low fuel loading, which results in fires of short duration that produce low to moderate subsurface heating. Semidesert rangeland fires typically create soil surface temperatures from 212 °F to 730 °F (100 °C to 388 °C). Soil organic matter begins changing chemically when heated to 200 °C and is completely destroyed at 450 °C (DeBano, 1990). Ambient soil surface temperature is generally higher after a fire due to the removal of vegetative cover and litter and to the enhanced thermal characteristics of charred material on the soil surface. Heating may cause changes in some physical properties of soils, as well, including the loss or reduction of structure, reduction of porosity, and alteration of color (Baker, 1990).
Frequent fires are presumed to deplete soil nutrients by volatilization and erosion. Biggs and others (n.d.) examined the effects of fire frequency on soil nutrients at three sites on the gunnery ranges of Fort Huachuca in southeastern Arizona. Due to differences in localized geography and climate and the absence of cattle grazing during recent decades, the findings from this study may not be directly applicable to the A-7 Ranch. However, the conclusions are worth noting in respect to changes in desert soil conditions subjected to fire.
Biggs and others (n.d.) measured the nutrient composition of soil and vegetation on one unburned site and two burned sites averaging three fires per decade and five fires per decade. The unburned site had an abundance of mature velvet mesquite (Prosopis velutina) and the unburned plots consisted primarily of desert grass species with widely scattered mesquite trees. Surface soil in the infrequently-burned site (two fires per decade) had higher average levels of total organic carbon, PO4, and NO3 than both the unburned and frequently-burned site (five fires per decade), although the highest concentrations of total organic carbon (TOC), nitrogen (N), and phosphorus (P) accumulated in soils beneath mesquite canopies. Mesquite trees appear to alter soil organic matter pools by concentrating key nutrients in their biomass and recycling them back into the soil as leaf litter decomposes. Fire results in the both the volatilization of N and P and deposition of N and P onto the soil surface where it is susceptible to loss from erosion and leaching. Soil pH was shown to increase with higher fire frequency. The cation exchange capacity was approximately 50% greater on the burned sites than on the unburned site. Lower levels of soil Na, K, and Ca were measured on burned sites. The total biomass of grasses and other shrubs was greater on burned sites than on the unburned site. Plants on the burned sites had lower concentrations of N and P but higher levels of carbon than plants on the unburned site. Carbon to nitrogen ratios at all three sites indicated that N was not a limiting factor in plant growth. Production of grasses and other forage plants was greatest on the infrequently burned site. This study recommended that the interval of fire occurrence should be greater than five years to avoid significant losses of soil macronutrients. The study also suggests that fire frequency not exceeding two fires per decade may be advantageous for preservation and restoration of grasslands.
4.5.2. Impacts to Desert Vegetation
One of the most significant changes in the Sonoran desert landscape since the late 1800s has been the invasion of woody plants into semidesert grasslands. The spread of shrubs and trees from low-lying riparian areas into upland areas has been associated with overgrazing, rodent seed dispersal, fire suppression, and climatic conditions (Cox et al., 1993; Zwolinski, 1990). Fire is one management tool that has been used to reduce woody plants and increase herbaceous biomass on desert rangeland. Frequent fires are detrimental to shrub establishment, but are less likely to reduce stands of mature seed bearing shrubs and trees. Reestablishment of herbaceous vegetation after fire varies depending upon species and climate (Cox et al., 1990). Tobosa grass, sacaton, alkali sacaton, and mixed grama ranges have been demonstrated to respond positively to burning (Wright, 1990). Wright (1990) recommended prescribed fire for the control of burroweed, snakeweed, creosote bush, and young mesquite trees.
The principal factors influencing burning potential are fuel moisture, fine fuel accumulation, preburn grazing intensity, and climatic variables. Predicting ecosystem responses to fire disturbance is difficult due to the range of variables influencing potential responses. Vegetation types, weather conditions, and topography control the extent and spread of fires by influencing fuel continuities and loading (Zwolinski, 1990). Annual weather patterns control fuel conditions and fire ignitions on a seasonal time scale (Swetnam, 1990).
The effect of fire on vegetation is influenced by a variety of factors including precipitation patterns before and after a burn, fluctuating climatic cycles, the composition of the vegetation, and the time of the burn. Drought can interact with fire to influence vegetation in several ways. Drought patterns determine the amount of fuel available to carry fires and influence the postburn response of vegetation to burning. Burning during a drought when soil moisture is low, or during midsummer when plants are beginning seed production may delay postburn recovery (Zwolinski, 1990).
The impacts of grazing management before and after a fire have a dramatic effect on the response of vegetation to fire. Preburn rest from grazing is required on most desert shrublands and grasslands to allow the accumulation of enough fine fuel to carry the fire. Without sufficient fine fuels a patchy fire may occur or the fire may not be able to carry at all. More than one year of preburn rest from grazing may be required to obtain adequate fuel to carry fire, especially in areas with severely depleted understories (Connors, personal communication). Severely depleted sites may require several years of rest before burning in order for key plants to regain vigor and reproductive capabilities. Prescribed burns following two years of better than average plant growth will improve the potential for grasses to recover quickly after burning (Martin, 1983; Wright, 1990).
Following burning rest from livestock grazing is required to allow seedling establishment and new growth from root structures and rhizomes. Management of livestock on a burned area is most critical the first growing season after fire, particularly in plant communities of arid and semiarid regions (Cox et al., 1990). Grazing management following burning may significantly affect the degree of change in forage species' productivity and possibly in the composition of the postburn vegetation. If livestock have premature access to the burn the full benefits of the prescribed fire may not be realized (Cable, 1967). Burned areas should be rested until a good ground cover and a litter layer are present to provide soil and watershed protection. Generally, at least two growing seasons of rest are recommended to allow reestablishment of preferred species and to deter reinvasion of shrubs (Connors, personalcommunication). A two growing-season rest after a burn greatly speeds the recovery and improvement of grass species. The length of the period of postburn rest from livestock use depends on the growing season conditions, (i.e. temperature and precipitation) prior to and following burning. Below normal precipitation may delay vegetative recovery. Bunch grasses require lighter utilization rates and longer rest periods following burning than do rhizomatous or annual plant species (Cox et al., 1990).
Timing of Prescribed Burns
The timing of prescribed burns must be coordinated with weather conditions, the livestock grazing schedule, the availability and moisture content of fine fuels to carry fire, and management objectives. Differing opinions exist on whether summer or fall burning is more advantageous for improving forage in semidesert grasslands (Martin, 1983; Wright, 1990). A disadvantage of June fires is that they often kill high percentages of several annual grass species, as well as of shrubs (Cable, 1967). Early summer fires tend to damage annual grasses, which have not had the opportunity to produce seed. Perennial grass species that are still dormant during early summer fires have the potential to resprout later in the season, particularly during periods of sufficient summer rains (Zwolinski, 1990). Some rangeland managers prefer fall burning in southeastern Arizona because the quantity of herbaceous fuel is the greatest at that time and perennial grasses are mainly dormant. Fall burning following a 12-month restriction on grazing has been demonstrated to produce the most optimal results in reestablishing native and exotic grasses on the Santa Rita Ranch south of Tucson, Arizona (Martin, 1983).
Cox and others (1990) investigated the responses of buffelgrass, big sacaton, and Lehmann's lovegrass to seasonal burning. Experimental plots were burned in the spring, summer, fall, and winter. Results from this study were highly variable over the three-year period following burning. The recovery rates of these three grass species were more highly correlated to precipitation than to the season of burning. Cox and others (1990) concluded that burning during any season reduces the amount of green herbage available for livestock grazing on all three grass species for at least two years.
Fire's Effects on Selected Plant Species on the A-7 Ranch
Whitethorn acacia ( Acacia constricta Benth.) and Velvet mesquite (Prosopis velutina [Sw.] DC.)
Of particular concern to ranchers in southeastern Arizona is the increasing abundance of native shrubs, such as whitethorn acacia and velvet mesquite (Cox et al., 1993). The dominance of these species has led to a reduction in perennial grass densities and has accelerated soil erosion throughout southeastern Arizona (Cooke and Reeves, 1976, in Cox et al., 1993). Periodic burning could maintain a grassland aspect by repeatedly topkilling some mesquites and killing most small seedlings, but the rapid rate of regrowth of mesquite would necessitate burning every five or six years to keep the mesquites from reaching reproductive maturity (Martin, 1983). Cox and others (1990) state that "mesquite burned once may be totally defoliated, but plants resprout within 36 months. Therefore, multiple burning strategies to reduce mature mesquite populations and kill seedlings will be necessary if the semiarid grasslands are to return."
The reduction of mesquite by fire is dependent on the age and size of the mesquite and the intensity of the burn (see Figure
4-16, below, for an illustration of differential mortality). The buildup of fine fuels following one or two years of above average rainfall increases the potential mortality of mesquite seedlings to fire. The percentage of mesquite killed by fire drops with increased age and size of the tree. Glendening and Paulson (1956, in Bahre and Shelton, 1993) reported that a single fire killed 60% of mesquite trees less than 13 mm in diameter, 20% of trees 25-51 mm in diameter, and 11% of trees over 127 mm in diameter. Martin (1983) found that, in a pasture where cattle had not grazed for twelve months, fire topkilled most small mesquites, but that, within five years, regrowth of mesquite equaled or exceeded preburn levels. Schmutz and others (1985) suggest that for effective mesquite control it would be necessary to burn more frequently than every seven years, and that there would be a need for an abundance of fine fuels permitting high-temperature fires. Mortality and topkill of velvet mesquite were higher for June fires than for October, November or February fires (Glendening and Paulson, 1955, in Martin, 1983). Prescribed burning for the purpose of mesquite eradication may be more effective during the summer season if the quantity of fine fuels is high enough to ensure hot fires.
|
|
|
Figure 4-16. Burned mesquite showing a dead individual in the foreground and a burned but re-leafing individual in the background. |

Cox and others (1993) found that fire killed 100% of both velvet mesquite and whitethorn acacia seed located on the soil surface, but had no measurable effects on the germination of seeds planted at a depth of 2 cm. The results of this study suggest that wildfire was an important factor in limiting mesquite and whitethorn acacia distribution on former semidesert grasslands. Humphrey (1974, in Bahre and Shelton, 1993) posits that the increased frequency of fire in southwestern grasslands prior to 1880 reduced the opportunity of mesquite seedlings to survive long enough to become mature trees and set seed.
Burroweed (Isocoma tenuisecta Greene) and Snakeweed (Gutierrezia microcephala [DC.] Gray)
Martin (1983) suggests that the effects of fall burning on perennial vegetation are only temporary. Shrubs susceptible to fire such as burroweed and snakeweed may be significantly reduced for one or two years, but these species may be expected to return to preburn densities within a few years (Cable, 1967; Martin, 1983; Schmutz et al., 1985). Schmutz and others (1985) reported that snakeweed crown cover seven years after fire was only slightly less on a burned area than on an unburned area. Following two prescribed burns in 1952 and 1955, the density of burroweed by 1965 was nearly double that of preburn levels (Cable, 1967). Cable (1967) found that burroweed density after fire was dependent on winter precipitation - increasing during years of high winter precipitation and declining during years of low winter precipitation. Burroweed seedlings quickly reinvade burned plots following above average winter precipitation. Schmutz and others (1985) attributed the rapid recovery of burroweed to a reduction in grass competition and to an increase in fertility from ash deposited on soil after fire. Martin's (1983) findings were inconclusive as to whether or not new burroweed plants had grown from seeds that survived the fires or from seeds imported from adjacent unburned plots. Mortality of burroweed in June fires was about twice as high as for fires in late fall or early spring (Tschirley and Martin, 1961, in Martin, 1983).
Arizona cottontop grass (Digitaria californica [Benth.] Henrard)
Schmutz and others (1985) found the recovery of Arizona cottontop to be slow under below-normal summer moisture conditions, reaching only approximately 10% of preburn levels seven years after burning. Martin (1983) reported that after burning, Arizona cottontop was rapidly reestablished on experimental plots following two years of above average winter and summer precipitation.
Black grama grass (Bouteloua eriopoda Torr.)
Burning has been found to severely reduce black grama on southwestern semidesert ranges (Cable, 1967). The poor reseeding potential of black grama in addition to its susceptibility to heavy grazing following burning limits its reestablishment (Martin, 1983). Wright (1990) recommends the avoidance of prescribed fire on rangeland with high concentrations of black grama. Three or four years of rest may be required before producing sufficient regrowth of black grama for grazing.
Lehmann's lovegrass (Eragrostis lehmanniana Nees.)
A positive-feedback pattern exists between Lehmann's lovegrass invasion and fire (see Figure 4-17, below). In areas where
Lehmann's lovegrass is present its dominance over native Arizona grasses has been positively correlated with rangeland fire (Ruyle et al., 1988; Anable et al., 1992). Martin (1983) found that the density of Lehmann's lovegrass increased to above preburn levels within three years following burning. Lehmann's lovegrass produces greater quantities of aboveground biomass than
native grasses (Cox et al., 1990). Lehmann's lovegrass also becomes established more readily and outcompetes native grasses after fire. These tendencies, in turn, result in greater quantities of fine fuels and increased natural and human-induced fire potential (Anable et al., 1992). Prescribed burns on rangeland containing Lehmann's lovegrass would likely further promote its spread and increased density.
|
|
|
Figure 4-17. Burned bunchgrass showing resprouting from the roots after a fire that was sufficient to kill nearby manzanita. |

Buffelgrass (Pennisetum ciliare [L.] Link )
Cox et al. (1990) recorded rapid recovery rates of buffelgrass after a summer fire, which was followed by above average summer precipitation. Buffelgrass yielded 25% more green herbage than unburned areas three months after the fire. During the following year, which received approximately 50% less summer rainfall, green herbage from the unburned area was equal to that of the burned area.
Prickly Pear and Cholla cacti (Opuntia spp.) and Barrel cactus (Ferocactus spp.)
Cactus has been shown to be adversely affected by repeated burning. Singing off cactus spines increases its palatability to both cattle and wildlife (Martin, 1983; Bock and Bock, 1990). Barrel cactus is particularly susceptible to fire (Wright, 1990). Prescribed burns at intervals of five or six years would most likely prevent encroachment by prickly pear and cholla, yet could simultaneously negatively impact other cactus species.
4.5.3. Fire's Effects: Summary
The response of vegetation to burning is variable and highly dependent on postburn precipitation. Using fire on the A-7 as a
mechanism to reduce the invasion of woody and undesirable range vegetation may be effective if applied at a frequency of one burn every five years. Recommended pre and postburn rest would restrict grazing for a minimum of two years under optimal precipitation conditions. More often, three years of rest would be required for sustained improvement of forage grasses. In a
one-burn-per-five years scenario would thus only permit two years of grazing. Cox and others (1990) conducted studies on 150 controlled burns in semiarid rangelands of the southwest over a 10-year period. In their final analysis they state, "using fire to improve the forage quality in semiarid deserts is difficult to justify when quality improvements are short-lived and herbage production is adversely affected for two or three years." The use of prescribed burns to increase forage in a grass banking system is limited and the utility of this technique in controlling woody plant species is debated. Reducing the A-7 herd size could improve the potential of prescribed burns for vegetation management.
4.6. HERBIVORY AND RANGE MANAGEMENT IN THE SEMIDESERT GRASSLAND-OAK SAVANNA ECOSYSTEM: CONCLUDING THOUGHTS ABOUT GRASS BANKING
Rangelands have been described as "managed natural pastures" (Hodgkinson & Mott, 1987:172), an epithet that incorporates at once the human, production, and consumption aspects of a grazing system (Figure 4-18). These components - people, grass, and cows - are subject to variability in the greater physical environment - to climate - and influence rangeland ecology in turn through disturbance of soils, hydrology, and vegetation from former, ungrazed states.
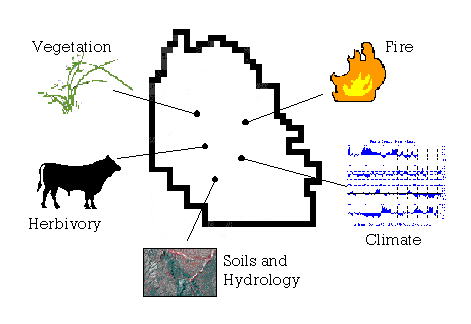
Figure 4-18. Variable aspects of the physical environment influencing management strategies on the A-7.
In uncertain environments, particularly where precipitation exhibits increased variability and soil moisture is a limiting factor to plant productivity, "the forage supply may vary considerably within and between years... Under these conditions, animal management is critical to the maintenance and productivity of the pasture base. Unfortunately, economic and logistic factors make it difficult to vary stock numbers in synchrony. Thus, the forage varies from being underused in good rainfall years to being overused in poor years, potentially causing range degradation" (Hodgkinson & Mott 1987:185). Management "aimed at maximizing animal productivity" (Hodgkinson & Mott, 1987:172) has in many cases resulted in irreversible change to semidesert grassland landscapes. Management in a "grass banking" system, coupled with suggestions to reduce stocking rates and to install more water sources and fencelines to facilitate pasture rotation, attempts to provide a means of accommodating variability while implementing a more sustainable approach to utilizing the forage resource. Part of such a system - formalized grass bank or not - will require, however, a shift in thinking about the economics of ranching from the short term attempt to beat the variability odds, to a long-term outlook that regards maintaining the land and its productivity as an investment with more predictable returns and something left to show for the effort (a legacy of maintenance, or improvement?) at the end.
4.7. REFERENCES CITED
Agnew, A. D. Q. (1997) Switches, pulses and grazing in arid vegetation. Journal of Arid Environments 37: 609-617.
Anable, M. E., M. P. McClaran, and G. B. Ruyle (1992) Spread of introduced Lehmann's lovegrass Eragrostis lehmanniana Nees. in southern Arizona, USA. Biological Conservation 61 (3): 181-188.
Bahre, C. J. (1995) Chapter 8: Human impacts on the grasslands of southeastern Arizona. In The Desert Grassland, edited by M. P. McClaran and T. R. Van Devender, pp. 230-264. University of Arizona Press, Tucson.
Bahre, C. J. (1996) A Legacy of Change: Historic human impact on vegetation in the Arizona borderlands. University of Arizona Press, Tucson.
Bahre, C. J. and M. L. Shelton (1993) Historic vegetation change, mesquite increases, and climate in southeastern Arizona. Journal of Biogeography 20 (5): 489-504.
Baker, M. B. (1990) Hydrologic and water quality effects of fire. General Technical Report RM-191. US Department of Agriculture, Forest Service, pp. 31-44.
Barry, R.G., and R.J. Chorley (1998) Atmosphere, Weather and Climate. Routledge, New York.
Biggs, T. H., R. H. Webb, and J. Quade (n.d.) Fire frequency and spatial variability of soil biogeochemistry and plant biochemistry in a southeastern Arizona desert grassland. Unpublished manuscript.
Bock, C. E. and J. H. Bock (1990) Effects of fire on wildlife in southwestern lowland habitats. General Technical Report RM-191. US Department of Agriculture, Forest Service, pp. 50-64.
Brady, W. (1995) 2.4 - Ecological Risks Facing Arizona's Grasslands. Arizona Comparative Environmental Risk Project (ACERP) Ecosystems Technical Committee Reports, Arizona Department of Environmental Quality, Phoenix.
Brown, D. E. (1994) Part 4 - Grasslands: Warm-Temperate Grasslands, 143.1-Semidesert Grassland, in Biotic Communities: Southwestern United States and Northwestern Mexico, edited by D. E. Brown, pp.122-131. University of Utah Press, Salt Lake City.
Brown, D. E. and C. H. Lowe (1994) Biotic Communities of the Southwest (Map). University of Utah Press, Salt Lake City.
Brown, P.W. (1999) Winter precipitation update. North Central Arizona Newsletter (Oct-Nov-Dec). University of Arizona, Tucson.
Buffington, L. C. and C. H. Herbel (1965) Vegetational Changes on a Semidesert Grassland Range from 1858 to 1963. Ecological Monographs 35 (2): 139-164.
Burgess, T. L. (1995) Desert Grassland, Mixed Shrub Savanna, Shrub Steppe, or Semidesert Scrub? The Dilemma of Coexisting Growth Forms. In The Desert Grassland, edited by M. P. McClaran and T. R. Van Devender, pp.31-67. University of Arizona Press, Tucson.
Cable, D. R. (1967) Fire effects on semidesert grasses and shrubs. Journal of Range Management 20: 170-176.
CDC (Climate Diagnostic Center-NOAA) SOI ranked by year 1896-1995 - La Niña and El Niño Extreme Years. Internet source: http://www.cdc.noaa.gov/Climaterisks/years.risk.html
CDC (Climate Diagnostic Center-NOAA) (2000) Multivariate ENSO Index. Internet source: http://www.cdc.noaa.gov/ENSO.mei_index.html
CLIMVIS(a) (Climatic Visualization System) - Global Historical Climatology Network. Internet source: <http://www5.ncdc.noaa.gov/cgi-bin/climvis/ghcn/tempV2.sh> via http://www.ncdc.noaa.gov/ol/climate/climatedata.html
CLIMVIS(b) (Climatic Visualization System) - Global Historical Climatology Network, National Climatic Data Center (NCDC). Internet source: <http://www5.ncdc.noaa.gov/cgi-bin/ghcn/precip.ghcncgi> via http://www.ncdc.noaa.gov/ol/climate/climatedata.html
CLIMVIS(c) (Climatic Visualization System) - Global Historical Climatological Network, National Climatic Data Center (NCDC). Climate Division Data: Arizona, section 7: Temperature, Precipitation, PDSI, PDHI data. Internet source: http://www.ncdc.noaa.gov/onlineprod/drought/xmgrg3.html
Connors, T. (2000) Personal communication - telephone conversation, 27 April 2000. USDA Forest Service, Coronado National Forest, Fire Management Division.
Cox, J. R., A. de Alba-Avila, R. W. Rice, and J. N. Cox (1993) Biological and physical factors influencing Acacia constricta and Prosopis velutina establishment in the Sonoran Desert. Journal of Range Management 46 (1): 43-48.
Cox, J. R., F. A. Ibarra, and M. H. Martin (1990) Fire effects on grasses in semiarid deserts. General Technical Report RM-191. US Department of Agriculture, Forest Service, pp. 43-49.
CPC (Climate Prediction Center-NOAA) (2000a) Weekly Palmer drought and crop moisture Data Products Explanation. Internet source: http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/cdus/palmer_drought/wpdanote.html
CPC (Climate Prediction Center-NOAA) (2000b) 102-year (1895-1996) Normal vs. ENSO-average precipitation (inches) by Climate Division. Internet source: http://www.cpc.ncep.noaa.gov/products/predictions/threats2/enso/elnino/az_bar.html
CPC (Climate Prediction Center-NOAA) (2000c) Cold and warm episodes by season. Internet source: http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.html
CPC (Climatic Prediction Center-NOAA) (1997) The 1997 El Niño: Potential effects in Arizona- November 1997 to April 1998. Internet source: http://www.cpc.ncep.noaa.gov/products/predictions/threats/enso/elnino/az.disc.html
Davey, J.R. (1980) Energy Flow Dynamics of a Grassland Ecosystem. Ph.D. Dissertation, Northern Arizona University, Flagstaff.
DeBano, L. F. (1990) Effects of fire on the soil resource in Arizona chaparral. General Technical Report RM-191. US Department of Agriculture, Forest Service, pp. 65-77.
Drought Information Center, NOAA (2000). The Palmer drought severity index. Internet source: http://www.drought.noaa.gov/palmer.html
EPA (Environmental Protection Agency) (1998) Climate change and Arizona. EPA 236-F-98-007c, September 1998.
Green, P. M., D. M. Legler, C. J. Miranda, and J. J. O'Brien (2000) The North American climate patterns associated with the El Niño-Southern Oscillation. Center for Ocean-Atmospheric Prediction Studies. Tallahassee, Florida. Florida State University. Internet source: http://www.coaps.fsu.edu/lib/booklet
Haile, A. (1981) Dynamics of Aboveground Net Primary Production in a Desert Grassland of Arizona. Ph.D. Dissertation, University of Arizona, Tucson.
Haskell, H. S. (1945) Effects of Conservative Grazing on a Desert Grassland Range as Shown by Vegetational Analysis. M.S. Thesis, University of Arizona, Tucson.
Hobbs, R. J. (1999) Gap vegetation map. NBII-USGS Biological Resources Division. Internet source: http://usgsbrd.srnr.arizona.edu.nbii
Hodgkinson, K. C. and J. J. Mott (1987) On Coping with Grazing. In Grazing-Lands Research at the Plant-Animal Interface, edited by F. P. Horn, et al., pp. 171-19?. Winrock International Institute, Morrilton, Arkansas.
Johnson, M. D. (1979) An analysis of spatial homogeneity and climatic change in Arizona summer rainfall. Thesis. Arizona State University, Tempe.
Kincaid, D. R. (1959) The Upper Desert Grassland of Southern Arizona - A Basic Ecological Analysis. M.S. Thesis, University of Arizona, Tucson.
Leopold, A. (1924) Grass, brush, timber, and fire in southern Arizona. Journal of Forestry 22: 1-10.
Malusa, J. and J. M. Porter (1990) A Biotic Survey of Buehman, Espiritu, Youtcy and Roble Canyons in the Redington Pass Region, Arizona. Unpublished manuscript on-file with Coronado N.F., Tucson.
Martin, S. C. (1983) Responses of semi-desert grasses and shrubs to fall burning. Journal of Range Management 36: 604-610.
McClaran, M. P. (1995) Desert Grasslands and Grasses. In The Desert Grassland, edited by M. P. McClaran and T. R. Van Devender, pp.1-30. University of Arizona Press, Tucson.
Naruk, S. J. and A. Bykerk-Kauffman (1990) Late Cretaceous and Tertiary Deformation of the Santa Catalina Metamorphic Core Complex, Arizona. In Geologic Excursions through the Sonoran Desert Region, Arizona and Sonora, edited by G. E. Gehrels and J. E. Spencer, pp.41-50. Special Paper 7, Arizona Geological Survey, Tucson.
NRCS (Natural Resources Conservation Service)-USDA (1999) Ecological Site Guides: Major Land Resource Area Descriptions, Area 41 - Southeastern Arizona Basin and Range. Internet source: http://az.nrcs.usda.gov/fotg/sec1/41.htm
Pagano, T. (1999) Southwest climate in a nutshell. Department of Hydrology and Water Resources, University of Arizona, Tucson.
Quimby, C. (1976) Bellota Range Environmental Analysis. United States Forest Service, Coronado N.F., Tucson.
Ruyle, G. B., B. A. Roundy, and J. R. Cox (1988) Effects of burning on germinability of Lehmann's lovegrass. Journal of Range Management 41: 404-406.
Schloderer, J. P. (1974) Geology and Kinematic Analysis of Deformation in the Redington Pass Area, Pima County, Arizona. M.S. Thesis, University of Arizona, Tucson.
Schmutz, E. M., K. M. Sourabie, and D. A. Smith (1985) The Page Ranch story - its vegetative history and management implications. Desert Plants 7 (1): 13-23.
Sellers, W. D., and R. H. Hill (1974) Arizona Climate: 1931-1972 (2nd edition). University of Arizona Press, Tucson.
Sheppard, P. R., A. C. Comrie, G. D. Packin, K. Angersbach, and M. K. Hughes (1999) The Climate of the Southwest (Draft). Internet source: http://tree.ltrr.arizona.edu/~sheppard/swclimate/swclim.htm
Simanton, J. R., G. D. Wingate, and M. A. Weltz (1990) Runoff and sediment from a burned sagebrush community. General Technical Report RM-191, US Department of Agriculture, Forest Service, pp.180-185.
Steila, D. (1972) Drought in Arizona: A Drought Identification Methodology and Analysis. Thesis. University of Arizona, Tucson.
Swetnam, T. W. (1990) Fire history and climate in the southwestern United States. General Technical Report RM-191. US Department of Agriculture, Forest Service, pp.6-17.
Swetnam, T. W. and J. L. Betancourt (1997) Mesoscale Ecological Responses to Climatic Variability in the American Southwest. Internet source: http://geochange.er.usgs.gov/sw/impacts/biology/fires_SOI/
USGS (U.S. Geological Survey) (2000) San Pedro River Nr. Redington, Arizona; Station Number: 09472000. Internet source: http://waterdata.usgs.gov/nwis-w/AZ/?ststnum=09472000
Woodhouse, C. A. (1997) Winter climate and atmospheric circulation patterns in the Sonoran Desert region, USA. International Journal of Climatology 17: 859-873.
Wright, H. A. (1990) Role of fire in the management of southwestern ecosystems. General Technical Report RM-191. US Department of Agriculture, Forest Service, pp.1-5.
Zimmermann, R. C. (1969) Plant Ecology of an Arid Basin, Tres Alamos-Redington Area, Southeastern Arizona. U.S. Geological Survey Professional Paper 485-D. U.S. Government Printing Office, Washington, D.C.
Zwolinski, M. J. (1990) Fire effects on vegetation and succession. General Technical Report RM-191, US Department of Agriculture, Forest Service, pp.19-24.
http://aria.arizona.edu/courses/arl642/grassbank2000/climate.html
Page last updated 07 February 2001